Энергетика мышечного сокращения. Инергию для работы мышц мы получаем в результате
Читайте также
А)только животных
В) только растений
С) только грибов
D) всех живых организмов
2) Получение энергии для жизнедеятельности организма происходит в результате:
A) размножения
B) дыхания
C) выделения
D) роста
3) Для большинства растений, птиц, зверей средой обитания является:
A) наземно-воздушная
B) водная
C) другой организм
D) почвенная
4) Цветки, семена и плоды характерны для:
A) хвойных растений
B) цветковых растений
C) плаунов
D) папоротников
5) Животные могут размножаться:
A) спорами
B) вегетативно
C) половым способом
D) делением клетки
6) Для того, чтобы не отравиться нужно собирать:
A) молодые съедобные грибы
B) грибы вдоль автомобильных дорог
C) ядовитые грибы
D) съедобные переросшие грибы
7) Запас минеральных веществ в почве и воде пополняется за счет жизнедеятельности:
А) производителей
B) разрушителей
C) потребителей
D) все ответы верны
8) Бледная поганка:
A) создает органические вещества на свету
B) переваривает питательные вещества в пищеварительной системе
C) всасывает питательные вещества гифами
D) захватывает питательные вещества ложноножками
9) Вставьте звено в цепь питания, выбрав из предложенных:
Овес- мышь- пустельга- .......
A) ястреб
B) чина луговая
C) дождевой червь
D) ласточка
10) Способность организмов реагировать на изменения окружающей среды называется:
A) выделение
B) раздражимость
C) развитие
D) обмен веществ
11) На среду обитания живых организмов воздействуют факторы:
A) неживой природы
B) живой природы
C) деятельность человека
D) все перечисленные факторы
12) Отсутствие корня характерно для:
A) хвойных растений
B) цветковых растений
C) мхов
D) папоротников
13) Тело протистов не может:
A) быть одноклеточным
B) быть многоклеточным
C) иметь органы
D) нет верного ответа
14) В результате фотосинтеза в хлоропластах спирогиры образуется (ются):
A) углекислый газ
B) вода
C) минеральные соли
D) нет верного ответа
1. выростами мембраны
2. полужидкой коллоидной капсулой
3. мезосомой
4. ядерным аппаратом
2. В хлоропластах формируются:
1. белки;
2. углеводы;
3. нуклеиновые кислоты;
4. жиры.
3. Лимфатические протоки впадают в:
1. правое предсердие;
2. аорту;
3. полые вены;
4. воротную вену печени.
4. Кристы – это образования:
1. мембраны ядра;
2. слепые ветви ЭПС;
3. мембраны лизосом;
4. внутренней мембраны митохондрий.
5. Биомасса – это:
1. вес тела;
2. плотность тела;
3. критерий биогеоценоза;
4. объем тела.
6. В одной семье 4 детей, имеющих I, II, III и IV группы крови. Родители имели группу крови:
1. I и IV;
2. II и III;
3. II и IV;
4. I и II;
7. Парные гены, расположенные в гомологичных хромосомах, контролирующие проявле-ние одного и того же признака, называют:
1. аллельными;
2. доминантными;
3. рецессивными;
4. сцепленными.
8. Фаза митоза, при котором хромосомы переместились в середину клетки.
1. Профаза
2. Метафаза
3. Анафаза
4. Телофаза
9. Что является примером пиноцитоза клетки?
1. Ускорение биохимических реакций
2. Выброс наружу пищевых остатков у инфузорий
3. Нагноение раны
4. Поступление воды в клетку
10. Конъюгация хромосом характерна для...
1. Процесса оплодотворения
2. Профазы 2-го деления мейоза
3. Митоза
4. Профазы 1-го деления мейоза
11. Клевер - это растение:
1. семейства сложноцветных, имеющее соцветие корзинку и плод семянку;
2. семейства бобовых, имеющее соцветие корзинку и плод семянку;
3. семейства бобовых, имеющее соцветие головку и плод боб;
4. семейства сложноцветных, имеющее соцветие головку и плод орешек.
12. Какой органоид обеспечивает транспорт веществ в клетке?
1. хлоропласт
2. митохондрия
3. рибосома
4. эндоплазматическая сеть
13. Что характерно для соматических клеток позвоночных животных?
1. имеют диплоидный набор хромосом
2. при слиянии образуют зиготу
3. участвуют в половом размножении
4. имеют одинаковую форму
14. Из споры папоротника развивается
1. заросток
2. корневище
3. стебель
4. основания корневищ
15. У человека на ранней стадии эмбрионального развития формируется...
1. Над хордой нервная трубка, а под ней кишечник
2. Над хордой кишечник и нервная трубка
3. Над хордой кишечник, а под ней нервная трубка
4. Под хордой кишечник и нервная трубка
16. Защитную роль по обеззараживанию веществ и заживлению ран в ротовой полости выполняет...
1. Лизоцим
2. Муцин
3. Амилаза
4. Мальтаза
17. Отношения, при которых присутствие одного вида исключает пребывание другого вида, – это...
1. Антогонизм
2. Конкуренция
3. Агрессия
4. Комменсализм
18. Какая группа тканей обладает свойствами возбудимости и сократимости:
1. мышечная;
2. эпителиальная;
3. нервная;
4. соединительная.
19. Найдите соответствие между компонентами желудочного сока и их функциями:
а) вода 1) создает среду и уничтожает микроорганизмов
б) соляная кислота 2) защищает стенки желудка
в) ферменты 3) растворяет вещества
г) слизь 4) расщепляют белки
1. а-3, б-1, в-4, г-2
2. а-4, б-1, в-3, г-2
3. а-3, б-1, в-2, г-4
4. а-3, б-2, в-4, г-1
20. Энергия необходимая для работы мышц, освобождается в процессе:
1. биосинтеза;
2. пищеварения;
3. распада органических веществ;
4. газообмена.
Задание 2.
Задание включает 2 теста, к каждому из них предложено по 6 вариантов отве-тов. Вам необходимо выбрать только три ответа.
1. Саморегуляция в экосистеме тайги проявляется в том, что:
1. численность деревьев сокращается в результате лесного пожара;
2. волки ограничивают рост численности кабанов;
3. массовое размножение короедов приводит к гибели деревьев;
4. численность белок зависит от урожая семян ели;
5. популяция кабанов полностью уничтожается волками;
6. совы и лисицы ограничивают рост численности мышей.
2. Какие процессы происходят в профазе первого деления мейоза?
1. образование двух ядер
2. расхождение гомологичных хромосом
3. образование метафазной пластинки
4. сближение гомологичных хромосом
5. обмен участками гомологичных хромосом
6. спирализация хромосом
Задание 3.
Дайте полный ответ
1. Почему под действием молочнокислых бактерий молоко, квашеная капуста приоб-ретают кислый вкус?
2. Могут ли белые кролики быть нечистопородными (гетерозиготными) по окраске шерсти?
Сокращение – изменение механического состояния миофибриллярного
сократительного аппарата мышечных волокон в результате дейтвия нервных
импульсов.
Скелетная мышца представляет собой сложную систему, преобразующую химическую
энергию в механическую работу и тепло.
По теории скольжения, в основе сокращения лежит механическое взаимодействие между миозиновыми и актиновыми миофиламентами благодоря образованию между ними в период активности попереречных мостиков.
Непосредственным источником энергии для мышечного сокращения является расщепление высокоэнергетического вещества АТФ. В мышце происходит также промежуточная реакция, вовлекающая 2-ое высокоэнергетическое вещество – креатинфосфат (КФ). Оно не может действовать как непосредственный источник энергии, поскольку его расщепление не оказывает влияние на сократительные белки мышцы. КФ обеспечивает энергией ресинтез АТФ. В свою очередь, энергия для ресинтеза КФ обеспечивается окислением.
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий на мембране в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазматического ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения.
Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция из саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют "электрическим сопряжением". Энергия гребкового движения одного мостика производит перемещение на 1% длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са2+-. Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФ-азы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению имеющихся мостиков и образованию в присутствии Са2+новых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом.
Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна - через 20 мс.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Основа сокращения мышцы - биохимические процессы, которые совершаются в 2 фазы: первую, анаэробную (бескислородную), и вторую, аэробную (кислородную). В каждой из этих фаз происходит расщепление веществ с освобождением энергии и их восстановление (ресинтез). Поэтому мышца, лишенная кислорода, может долго работать при условии удаления остаточных продуктов обмена веществ. Следует учесть, что исключительная роль в биохимических процессах первой фазы принадлежит кислоторастворимым фосфорным соединениям, относительное содержание которых в скелетных мышцах кролика равняется (в процентах кислоторастворимого фосфора): аденозинтрифосфорная кислота (АТФ) - 35, креатинфосфорная кислота - 45, гексозомонофосфорная кислота - 5, неорганическая фосфорная кислота - 10.
Первое место в анаэробной фазе химических процессов занимает расщепление АТФ, которое происходит в первую очередь и с наибольшей скоростью. АТФ дефосфорилируется благодаря ферментативному действию мышечных белков. Отщепляя группу фосфорной кислоты, АТФ превращается сначала в аденозиидифосфорную (АДФ), а затем при потере еще одной группы фосфорной кислоты - в адениловую кислоту. При отщеплении каждого моля фосфорной кислоты освобождается 33,5 - 50,2 кДж, а в результате отщепления двух молей фосфорной кислоты - до 100,5 кДж при превращении каждого моля АТФ в адениловую кислоту. Эта энергия используется для сокращения мышц. Так как АТФ расходуется, то длительная мышечная работа невозможна без ресинтеза АТФ.
Ресинтез АТФ в анаэробных условиях происходит за счет энергии, которая освобождается при втором, более медленном процессе дефосфорилироваиия креатинфосфорной кислоты на креатин и фосфорную кислоту. При этом расщеплении на каждый моль креатинфосфата освобождается 46 кДж. Часть креатина необратимо распадается.
И, наконец, происходит третий, наиболее медленный процесс анаэробной фазы -
расщепление гексозофосфорной кислоты на фосфорную и молочную кислоты. При этом
процессе на каждый моль образовавшейся молочной кислоты выделяется 104,6 кДж
Гликоген, присоединяя фосфорную кислоту, сначала превращается в
гексозомонофосфат, а затем в гексозодифосфат с небольшим поглощением энергии.
Гексозодифосфат распадается на молочную и фосфорную кислоты с значительным
освобождением энергии.
Ресинтез АТФ совершается в течение тысячных долей секунды в результате присоединения к адениловой кислоте групп фосфорной кислоты, освободившихся при расщеплении креатинфосфорной кислоты и гексозофосфата, Ресинтез креатинфосфорной кислоты происходит в результате присоединения к креатину фосфорной кислоты, освободившейся при расщеплении гексозофосфата. Энергия, освободившаяся при расщеплении креатинфосфорной кислоты и гексозофосфата, обеспечивает процессы ресинтеза, главным образом АТФ. Так как избыток адениловой кислоты, образовавшейся в первой фазе из АТФ, необратимо дезаминируется и превращается в инозиновую кислоту и аммиак, то при полном ресинтезе АТФ вначале идет ресинтез адениловой кислоты. Адениловая кислота ресинтезируется из инозиновой кислоты и аммиака, освобождающегося при дезаминировании аминокислот.
Во второй, аэробной, фазе биохимических процессов освобождается наибольшее
количество энергии, которое используется как для ресинтеза соединений,
расщепляющихся в первой и второй фазах, так и главным образом для мышечной
деятельности.
В результате гликолиза (бескислородного распада глюкозы на две частицы молочной
кислоты) и расщепления гексозофосфатов образуется молочная кислота, которая
окисляется до углекислоты и воды. Но не вся молочная кислота окисляется, а
примерно 76 часть (при утомлении около 1/4). При этом расщепление каждого моля
молочной кислоты освобождает 1465,5 кДж. За счет этой энергии происходит
ресинтез остальной части молочной кислоты (5/6 – 3/4) до глюкозы и гликогена, а
также ресинтез АТФ и креатинфосфорной кислоты.
Следовательно, наибольшее количество энергии при мышечной работе освобождается при окислении углеводов. Гликоген содержится главным образом в анизотропных дисках. В мышцах в покое, а также при восстановлении после сокращения кроме углеводов расщепляется небольшая часть белков и липидов. При достаточном кровообращении и содержании глюкозы и кислорода в крови мышца работает за счет энергии окислительных процессов. В покое мышца использует около 5% кислорода, поступающего с кровью. Ресинтез веществ, расщепляющихся при мышечной деятельности, увеличивается при повышении интенсивности тканевого дыхания. Таким образом, процесс расщепления усиливает ресинтез, что ведет к наиболее экономному использованию веществ и энергии.
В результате сокращения в мышечных волокнах возникает напряжение. Эта способность развивать напряжение является основным свойством мышцы. Напряжение, развиваемое мышцами при сокращении, реализуется по-разному. Если внешняя нагрузка меньше, чем напряжение сокращающейся мышцы, то мышца укорачивается и вызывает движение – это изотонический (концентрический, миометрический) тип сокращения. Поскольку в экспериментальных условиях при электрическом раздражении укорочение мышцы происходит при постоянном напряжении = внешней нагрузке.
Если внешняя нагрузка больше, чем напряжение, развиваемое мышцей во время
сокращения, то такая мышца растягивается при сокращении – это эксцентрический (плиометрический)
тип сокращения.
Изотонический и эксцентрический типы сокращения, т.е. сокращения. при которых
мышца изменяет свою длину, относятся к динамической форме сокращения.
Сокращение мышцы, при котором она развивает напряжение, но не изменяет своей длины, называется изометрическим. Это статическая форма сокращения. Она возникает в 2-ух случаях: либо когда внешняя нагрузка = напряжению, развиваемому мышцей при сокращении, либо когда внешняя нагрузка превышает напряжение мышцы, но отсутствуют условия для растяжения мышцы под влиянием этой внешней нагрузки.
При динамических формах сокращения производится внешняя работа – при изотоническом сокращении положительная, при эксцентрическом отрицательная. Величина работы в обоих случаях может быть определена как произведение внешней нагрузки (поднятого веса) на пройденное расстояние. При изометрическом сокращении "расстояние" = 0, и, согласно физическому закону, в этом случае мышца не производит никакой работы.
Если внешняя нагрузка на мышцы меньше, чем ее напряжение, мышца укорачивается и вызывает движение. Это концентрический, или миометрический, тип сокращения. В экспериментальных условиях при электрическом раздражении изолированной мышцы ее укорочение происходит при постоянном напряжении, равном внешней нагрузке. Поэтому этот тип сокращения называют также изотоническим (изос - равный, тонус - напряжение).
Если внешняя нагрузка на мышцы больше, чем ее напряжение, развиваемое во время сокращения, мышца растягивается (удлиняется). Это эксцентрический, или плиометрический, тип сокращения. Концентрический и эксцентрический типы сокращения, т.е. сокращения, при которых мышца изменяет длину, относятся к динамической форме сокращения.
Сокращение мышцы, при котором она развивает напряжение, но не изменяет своей длины, называется изометрическим (изос - равный, метр - длина). Это статическая форма сокращения. Она возникает в двух случаях: когда внешняя нагрузка равна напряжению, развиваемому мышцей при сокращении, либо когда внешняя нагрузка превышает напряжение мышцы, но отсутствуют условия для растяжения мышцы под влиянием этой внешней нагрузки.
При динамических формах сокращения производится внешняя работа: при концентрическом сокращении - положительная, при эксцентрическом - отрицательная. Величина работы в обоих случаях определяется как произведение внешней нагрузки (поднятого веса) на пройденное расстояние. При изометрическом сокращении "расстояние" равно нулю, и, согласно физическому закону, в этом случае мышца не производит никакой работы. Однако с физиологической точки зрения изометрическое сокращение требует расхода энергии и может быть очень утомительным. В этом случае работа может быть определена как произведение величины напряжения мышцы на время ее сокращения (т.е. эквивалентно импульсу силы в физике). Во время изометрического сокращения в тепло превращается вся выделяемая мышцей энергия, а динамическом сокращении не менее 50% ее энергии.
Энергетические резервы человека (с массой тела 75 кг)
Целая мышца содержит большое количество соединительной ткани; кроме того, саркомеры в разных частях мышцы не всегда сокращаются с одинаковой силой. В результате данная кривая по сравнению с кривой для отдельного мышечного волокна имеет в определенной степени иные размерности, но демонстрирует ту же основную форму наклона в области нормального диапазона сокращения.
На рисунке видно, что при длине мышцы, соответствующей ее состоянию покоя (т.е. когда длина саркомера около 2 мкм), активация мышцы приводит к ее сокращению, сила которого близка к максимальной. Однако увеличение напряжения, происходящее во время сокращения и называемое активным напряжением, снижается по мере растяжения мышцы за пределы ее нормальной длины (т.е. за пределы длины саркомера примерно в 2,2 мкм). Это демонстрируется на рисунке уменьшением длины стрелки при длине мышцы больше нормы.
Без нагрузки скелетная мышца сокращается чрезвычайно быстро, достигая максимума примерно за 0,1 сек для усредненной мышцы. При наличии нагрузки по мере ее возрастания скорость сокращения постепенно снижается. Когда нагрузка возрастает до величины, равной максимальной силе, которую способна развить мышца, скорость сокращения становится нулевой, и в результате укорочения мышцы нет, несмотря на ее активацию.
Снижение скорости сокращения при увеличении нагрузки связано с тем фактом, что нагрузка на сокращающуюся мышцу противостоит силе, развиваемой мышцей при сокращении. Следовательно, общая сила, доступная для развития скорости укорочения, соответственно уменьшается.
При сокращении против нагрузки мышца выполняет работу. Это значит, что энергия перемещается от мышцы к внешней нагрузке, чтобы поднять объект до большей высоты или преодолеть сопротивление движению.
В математическом выражении работа
определяется следующим уравнением:
W = L х D,
где W - производимая работа, L - нагрузка и D - расстояние движения против нагрузки. Источником энергии, необходимой для выполнения работы, являются химические реакции в мышечных клетках во время сокращения, которые приведены в следующих разделах.
Источники энергии для мышечного сокращения
Мы уже знаем, что мышечное сокращение зависит от энергии , поставляемой АТФ. Большая часть этой энергии тратится на приведение в действие храпового механизма, с помощью которого поперечные мостики тянут актиновые нити. Однако небольшое количество энергии необходимо: (1) для откачивания ионов кальция из саркоплазмы в саркоплазматический ретикулум после завершения сокращения; (2) на активное перемещение ионов натрия и калия через мембрану мышечного волокна с целью поддержания соответствующей ионной среды для распространения потенциалов действия по волокну.
Концентрация АТФ в мышечном волокне (примерно 4 ммоль/л) достаточна для поддержания максимального сокращения в течение не более 1-2 сек. АТФ расщепляется с формированием АДФ и выделением энергии, которая передается механизму сокращения мышечного волокна. В течение следующей доли секунды АДФ рефосфорилируется, формируя новую молекулу АТФ, что позволяет мышце продолжать свое сокращение. Существуют несколько источников энергии для восстановления АТФ.
Первым источником энергии , используемым для восстановления АТФ, является креатинфосфат - вещество с высокоэнергетической фосфатной связью, подобной связям АТФ. Высокоэнергетическая фосфатная связь креатинфосфата имеет несколько большее количество свободной энергии, чем каждая связь АТФ. Креатинфосфат немедленно расщепляется, и освобожденная энергия вызывает связывание нового фосфата с АДФ, восстанавливая АТФ. Однако общее количество креатинфосфата в мышечном волокне также очень незначительно - примерно в 5 раз больше, чем АТФ. Следовательно, общий запас энергии в виде АТФ и креатинфосфата в мышце способен обеспечить максимальное сокращение в течение лишь 5-8 сек.
Вторым важным источником энергии , используемым для восстановления как АТФ, так и креатинфосфата, является гликолиз гликогена, предварительно накопленного в мышечных клетках. Быстрое ферментативное разрушение гликогена до пировиноградной кислоты, а затем до молочной кислоты освобождает энергию, которая используется для превращения АДФ в АТФ. Затем АТФ может участвовать непосредственно в обеспечении энергией дополнительного мышечного сокращения, а также в восстановлении запасов креатинфосфата.
Механизм гликолиза имеет двойное значение. Во-первых, гликолитические реакции могут происходить даже при отсутствии кислорода, и мышечное сокращение может поддерживаться в течение нескольких секунд, а иногда более 1 мин, даже если доставка кислорода из крови не доступна. Во-вторых, скорость образования АТФ с помощью гликолиза примерно в 2,5 раза выше, чем при образовании АТФ в реакциях питательных веществ клетки с кислородом. Однако конечных продуктов гликолиза в мышечных клетках накапливается так много, что примерно через 1 мин гликолиз также теряет способность поддерживать максимальное мышечное сокращение.
Третьим и решающим источником энергии является окислительный метаболизм, т.е. комбинирование кислорода с конечными продуктами гликолиза и различными другими клеточными питательными веществами с освобождением энергии. Более 95% всей энергии, используемой мышцей для непрерывного длительного сокращения, извлекается из этого источника. Потребляемыми питательными веществами являются углеводы, жиры и белки. Для чрезвычайно длительной максимальной мышечной активности, продолжающейся в течение многих часов, основная часть энергии получается из жиров, но при мышечной активности длительностью 2-4 ч до половины энергии может получаться из накопленных углеводов.
Сокращение мышц — это сложный процесс, состоящий из целого ряда этапов. Главными составляющими здесь являются миозин, актин, тропонин, тропомиозин и актомиозин, а также ионы кальция и соединения, которые обеспечивают мышцы энергией. Рассмотрим виды и механизмы мышечного сокращения. Изучим, из каких этапов они состоят и что необходимо для цикличного процесса.
Мышцы
Мышцы объединяются в группы, у которых одинаковый механизм мышечных сокращений. По этому же признаку они и разделяются на 3 вида:
- поперечно-полосатые мышцы тела;
- поперечно-полосатые мышцы предсердий и сердечных желудочков;
- гладкие мышцы органов, сосудов и кожи.
Поперечно-полосатые мышцы входят в опорно-двигательный аппарат, являясь его частью, так как помимо них сюда входят сухожилия, связки, кости. Когда реализуется механизм мышечных сокращений, выполняются следующие задачи и функции:
- тело передвигается;
- части тела перемещаются друг относительно друга;
- тело поддерживается в пространстве;
- вырабатывается тепло;
- кора активируется посредством афферентации с рецептивных мышечных полей.
Из гладких мышц состоит:
- двигательный аппарат внутренних органов, в который входят легкие и пищеварительная трубка;
- лимфатическая и кровеносная системы;
- система мочеполовых органов.
Физиологические свойства

Как и у всех позвоночных животных, в человеческом организме выделяют три самых важных свойства волокон скелетных мышц:
- сократимость — сокращение и изменение напряжения при возбуждении;
- проводимость — движение потенциала по всему волокну;
- возбудимость — реагирование на раздражитель посредством изменения мембранного потенциала и ионной проницаемости.
Мышцы возбуждаются и начинают сокращаться от идущих от центров. Но в искусственных условиях используют тогда может раздражаться напрямую (прямое раздражение) или через нерв, иннервирующий мышцу (непрямое раздражение).
Виды сокращений
Механизм мышечных сокращений подразумевает преобразование химической энергии в механическую работу. Этот процесс можно измерить при эксперименте с лягушкой: ее икроножную мышцу нагружают небольшим весом, а затем раздражают легкими электроимпульсами. Сокращение, при котором мышца становится короче, называется изотоническим. При изометрическом сокращении укорачивания не происходит. Сухожилия не позволяют при развитии укорачиваться. Еще один ауксотонический механизм мышечных сокращений предполагает условия интенсивных нагрузок, когда мышца укорачивается минимальным образом, а сила развивается максимальная.
Структура и иннервация скелетных мышц
В поперечно-полосатые скелетные мышцы входит множество волокон, находящихся в соединительной ткани и крепящихся к сухожилиям. В одних мышцах волокна расположены параллельно длинной оси, а в других они имеют косой вид, прикрепляясь к центральному тяжу сухожильному и к перистому типу.

Главная особенность волокна заключается в саркоплазме массы тонких нитей — миофибрилл. В них входят светлые и темные участки, чередующиеся друг с другом, а у соседних поперечно-полосатые волокна находятся на одном уровне — на поперечном сечении. Благодаря этому получается поперечная полосатость по всему волокну мышц.
Саркомером является комплекс из темного и двух светлых дисков, и он отграничивается Z-образными линиями. Саркомеры — это сократительный аппарат мышцы. Получается, что сократительное мышечное волокно состоит из:
- сократительного аппарата (системы миофибрилл);
- трофического аппарата с митохондриями, комплексом Гольджи и слабой ;
- мембранного аппарата;
- опорного аппарата;
- нервного аппарата.
Мышечное волокно разделяется на 5 частей со своими структурами и функциями и является целостной частью ткани мышц.
Иннервация
Этот процесс у поперечно-полосатых мышечных волокон реализуется посредством нервных волокон, а именно аксонов мотонейронов спинного мозга и головного ствола. Один мотонейрон иннервирует несколько волокон мышц. Комплекс с мотонейроном и иннервируемыми мышечными волокнами называют нейромоторной (НМЕ), или (ДЕ). Среднее число волокон, которые иннервирует один мотонейрон, характеризует величину ДЕ мышцы, а обратную величину называют плотностью иннервации. Последняя является большой в тех мышцах, где движения небольшие и «тонкие» (глаза, пальцы, язык). Малое ее значение будет, напротив, в мышцах с «грубыми» движениями (например, туловище).
Иннервация может быть одиночной и множественной. В первом случае она реализуется компактными моторными окончаниями. Обычно это характерно для крупных мотонейронов. (называющиеся в этом случае физическими, или быстрыми) генерируют ПД (потенциалы действий), которые распространяются на них.
Множественная иннервация встречается, к примеру, во внешних глазных мышцах. Здесь не генерируется потенциал действия, так как в мембране нет электровозбудимых натриевых каналов. В них распространяется деполяризация по всему волокну из синаптических окончаний. Это необходимо для того, чтобы привести в действие механизм мышечного сокращения. Процесс здесь происходит не так быстро, как в первом случае. Поэтому его называют медленным.
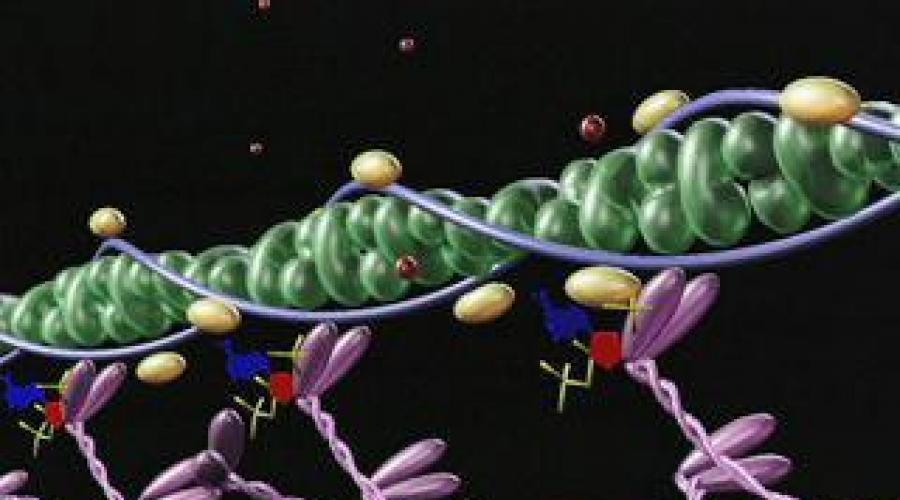
Структура миофибрилл
Исследования мышечного волокна сегодня проводятся на основе рентгеноструктурного анализа, электронной микроскопии, а также гистохимическими методами.
Рассчитано, что в каждую миофибриллу, диаметр которой составляет 1 мкм, входит примерно 2500 протофибрилл, то есть удлиненных полимеризованных молекул белков (актина и миозина). Актиновые протофибриллы в два раза тоньше миозиновых. В покое эти мышцы находятся так, что актиновые нити кончиками проникают в промежутки между миозиновыми протофибриллами.
Узкая светлая полоса в диске А свободна от актиновых нитей. А мембрана Z скрепляет их.
На миозиновых нитях есть поперечные выступы длиной до 20 нм, в головках которых находится порядка 150 молекул миозина. Они отходят биополярно, и каждая головка соединяет миозиновую с актиновой нитью. Когда происходит усилие актиновых центров на нитях миозина, актиновая нить приближается к центру саркомера. В конце миозиновые нити доходят до линии Z. Тогда они занимают собой весь саркомер, а актиновые находятся между ними. При этом длина диска I сокращается, а в конце он исчезает полностью, вместе с чем линия Z становится толще.
Так, по теории скользящих нитей, объясняется сокращение длины волокна мышцы. Теория, получившая название «зубчатого колеса», была разработана Хаксли и Хансоном в середине двадцатого века.

Механизм мышечного сокращения волокна
Главным в теории является то, что не нити (миозиновые и актиновые) укорачиваются. Длина их остается неизменной и при растяжении мышц. Но пучки тонких нитей, проскальзывая, выходят между толстыми нитями, уменьшается степень их перекрытия, таким образом происходит сокращение.
Молекулярный механизм мышечного сокращения посредством скольжения актиновых нитей заключается в следующем. Миозиновые головки соединяют протофибриллу с актиновой. При их наклонах происходит скольжение, двигающее актиновую нить к центру саркомера. За счет биполярной организации миозиновых молекул на обеих сторонах нитей создаются условия для скольжения актиновых нитей в разные стороны.
При расслаблении мышц миозиновая головка отходит от актиновых нитей. Благодаря легкому скольжению расслабленные мышцы растяжению сопротивляются гораздо меньше. Поэтому они пассивно удлиняются.
Этапы сокращения
Механизм мышечного сокращения кратко можно подразделить на следующие этапы:
- Мышечное волокно стимулируется, когда потенциал действия поступает от мотонейронов из синапсов.
- Потенциал действия создается на мембране мышечного волокна, а затем распространяется к миофибриллам.
- Совершается электромеханическое сопряжение, представляющее собой преобразование электрического ПД в механическое скольжение. В этом обязательно участвуют ионы кальция.
Ионы кальция

Для лучшего понимания процесса активации волокна ионами кальция удобно рассмотреть структуру актиновой нити. Длина ее составляет порядка 1 мкм, толщина — от 5 до 7 нм. Это пара закрученных ниток, которые напоминают мономер актина. Примерно через каждые 40 нм здесь находятся сферические тропониновые молекулы, а между цепями — тропомиозиновые.
Когда ионы кальция отсутствуют, то есть миофибриллы расслабляются, длинные тропомиозиновые молекулы блокируют крепление актиновых цепей и мостиков миозина. Но при активизации ионов кальция тропомиозиновые молекулы опускаются глубже, и участки открываются.
Тогда миозиновые мостики прикрепляются к актиновым нитям, а АТФ расщепляется, и сила мышц развивается. Это становится возможным за счет воздействия кальция на тропонин. При этом молекула последнего деформируется, проталкивая тем самым тропомиозин.
Когда мышца расслаблена, в ней на 1 грамм сырого веса содержится больше 1 мкмоль кальция. Соли кальция изолированы и находятся в особых хранилищах. В противном случае мышцы бы все время сокращались.
Хранение кальция происходит следующим образом. На разных участках мембраны клетки мышцы внутри волокна имеются трубки, через которые происходит соединение со средой вне клеток. Это система поперечных трубочек. А перпендикулярно ей находится система продольных, на концах которых — пузырьки (терминальные цистерны), расположенные в непосредственной близости к мембранам поперечной системы. Вместе получается триада. Именно в пузырьках хранится кальций.
Так ПД распространяется внутрь клетки, и происходит электромеханическое сопряжение. Возбуждение проникает в волокно, переходит в продольную систему, высвобождает кальций. Таким образом осуществляется механизм сокращения мышечного волокна.

3 процесса с АТФ
При взаимодействии обеих нитей при наличии ионов кальция немалая роль отводится АТФ. Когда реализуется механизм мышечного сокращения скелетной мышцы, энергия АТФ применяется для:
- работы насоса натрия и калия, который поддерживает постоянную концентрацию ионов;
- этих веществ по разные стороны мембраны;
- скольжения нитей, укорачивающих миофибриллы;
- работы насоса кальция, действующего для расслабления.
АТФ находится в клеточной мембране, нитях миозина и мембранах ретикулума саркоплазматического. Фермент расщепляется и утилизируется миозином.
Потребление АТФ
Известно, что миозиновые головки взаимодействуют с актином и содержат элементы для расщепления АТФ. Последняя активизируется актином и миозином при наличии ионов магния. Поэтому расщепление фермента происходит при прикреплении миозиновой головки к актину. При этом чем больше поперечных мостиков, тем скорость расщепления будет выше.
Механизм АТФ
После завершения движения молекула АФТ обеспечивает энергией для разделения участвующих в реакции миозина и актина. Миозиновые головки разделяются, АТФ расщепляется до фосфата и АДФ. В конце подсоединяется новая АТФ-молекула, и цикл возобновляется. Таковым является механизм мышечного сокращения и расслабления на молекулярном уровне.
Активность поперечных мостиков будет продолжаться лишь до тех пор, пока происходит гидролиз АТФ. При блокировке фермента мостики не станут снова прикрепляться.
С наступлением смерти организма уровень АТФ в клетках падает, и мостики остаются устойчиво прикрепленными к актиновой нити. Так происходит стадия трупного окоченения.

Ресинтез АТФ
Ресинтез возможно реализовать двумя путями.
Посредством ферментативного переноса от креатинфосфата фосфатной группы на АДФ. Так как запасов в клетке креатинфосфата намного больше АТФ, ресинтез реализуется очень быстро. В то же время посредством окисления пировиноградной и молочной кислот ресинтез будет осуществляться медленно.
АТФ и КФ могут исчезнуть полностью, если ресинтез будет нарушен ядами. Тогда и кальциевый насос прекратит работу, вследствие чего мышца необратимо сократится (то есть настанет контрактура). Таким образом, нарушится механизм мышечного сокращения.
Физиология процесса
Подытоживая вышесказанное, отметим, что сокращение волокна мышцы состоит в укорочении миофибрилл в каждом из саркомеров. Нити миозина (толстые) и актина (тонкие) связаны концами в расслабленном состоянии. Но они начинают скользящие движения друг навстречу к другу, когда реализуется механизм мышечного сокращения. Физиология (кратко) объясняет процесс, когда под влиянием миозина выделяется необходимая энергия для преобразования АТФ в АДФ. При этом активность миозина будет реализована лишь при достаточном содержании ионов кальция, накапливающихся в саркоплазматической сети.
Сокращение и напряжение мышцы осуществляется за счет энергии, освобождающейся при химических превращениях, которые происходят при поступлении в мышцу нервного импульса или нанесении на нее непосредственного раздражения. В качестве основного поставщика энергии выступает АТФ (аденозинтрифосфорная кислота).
АТФ в организме играет роль "универсальной валюты", идущей на оплату всех энергетических потребностей живых клеток. Так как запасы АТФ в мышцах невелики и, чтобы поддерживать их деятельность, необходим непрерывный ресинтез АТФ. Его восполнение и образование энергии в принципе происходит двумя способами - в зависимости от того присутствует при этом кислород или нет.
Реакции, совершающиеся в бескислородной среде получили название анаэробных. Освобождение энергии в этом случае происходит за счет мгновенного расщепления богатых энергией веществ на менее богатые. Последнее звено в этом расщеплении - когда гликоген превращается в молочную кислоту.
Гликоген - сложный вид сахара, родственный крахмалу. Сахар и другие виды углеводов, которые мы потребляем, накапливается в организме в виде гликогена. Следовательно, для простоты можно записать:
Этот механизм расщепления может давать большой эффект и он может использоваться при кратковременной максимальной работе (спринтерский бег, бег вверх по лестнице), когда необходимо внезапно проявить силу, а кровоснабжение мышц при этом недостаточно. Недостаток же заключается в том, что в работающих мышцах накапливается молочная кислота и им становится трудно справляться с воздействием кислой среды. Молочная кислота для мышцы является веществом утомления, и поэтому мышца может работать только незначительное время.
Реакции, происходящие с участием кислорода, получили название аэробных. Образование энергии и восстановление запасов АТФ в этом случае происходит за счет окисления углеводов и жиров. При этом образуются углекислый газ и вода. Часть энергии расходуется на восстановление молочной кислоты в глюкозу и гликоген. При этом обеспечивается ресинтез АТФ.
Аэробный ресинтез АТФ отличается высокой экономичностью, а также универсальностью в использовании субстратов: окисляются все органические вещества организма (аминокислоты, белки, углеводы, жирные кислоты и др.). Однако он требует потребления кислорода, доставка которого в мышечную ткань обеспечивается дыхательной и сердечно-сосудистой системами, что естественно связано с их напряжением. Кроме того развертывание аэробного образования АТФ продолжительно по времени и невелико по мощности.
58. Одиночное и тетанические сокращения
При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается одиночным сокращением. Эта форма механической реакции состоит из 3 фаз: латентного или скрытого периода, фазы сокращения и фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1.5-2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции - наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения - тетанус. Различают 2 формы тетануса - зубчатый тетанус, возникающий при более редком раздражении, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных одиночных сокращений, и сплошной или гладкий тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения.
Таким образом, (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5-8 имп. в 1с) возникают одиночные сокращения, при увеличении частоты (15-20 имп. в 1с) - зубчатый тетанус, при дальнейшем нарастании частоты (25-60 имп. в 1 с) - гладкий тетанус. Одиночное сокращение - более слабое и менее утомительное, чем титаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно поддерживая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверхдлинные дистанции). При этом отдельные ДЕ
могут развивать как одиночные, так и титанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т.е. одновременное возбуждение практически всех ДЕ. Это, в свою очередь, требует одновременной активации соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное титаническое сокращение.
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон "Все или ничего"). В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.